
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
Chains :
E.C.6.3.2.19
- Transferred entry: 2.3.2.23, 2.3.2.27 and 6.2.1.45.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + ubiquitin + protein lysine = AMP + diphosphate + protein N-ubiquityllysine
|
|
|
|
|
|
 ATP
ATP
|
+
|
ubiquitin
|
+
|
 protein lysine
protein lysine
|
=
|
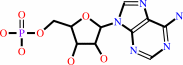 AMP
AMP
|
+
|
 diphosphate
diphosphate
|
+
|
protein N-ubiquityllysine
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Acta Crystallogr D Biol Crystallogr
62:1300-1310
(2006)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Crystal structures of two peptide-HLA-B*1501 complexes; structural characterization of the HLA-B62 supertype.
|
|
G.Røder,
T.Blicher,
S.Justesen,
B.Johannesen,
O.Kristensen,
J.Kastrup,
S.Buus,
M.Gajhede.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
MHC class I molecules govern human cytotoxic T cell responses. Their specificity
determines which peptides they sample from the intracellular protein environment
and then present to human cytotoxic T cells. More than 1100 different MHC class
I proteins have been found in human populations and it would be a major
undertaking to address each of these specificities individually. Based upon
their peptide binding specificity, they are currently subdivided into 12
supertypes. Several of these HLA supertypes have not yet been described at the
structural level. To support a comprehensive understanding of human immune
responses, the structure of at least one member of each supertype should be
determined. Here, the structures of two immunogenic peptide-HLA-B*1501 complexes
are described. The structure of HLA-B*1501 in complex with a peptide (LEKARGSTY,
corresponding to positions 274-282 in the Epstein-Barr virus nuclear antigen-3A)
was determined to 2.3 A resolution. The structure of HLA-B*1501 in complex with
a peptide (ILGPPGSVY) derived from human ubiquitin-conjugating enzyme-E2
corresponding to positions 91-99 was solved to 1.8 A resolution. Mutual
comparisons of these two structures with structures from other HLA supertypes
define and explain the specificity of the P2 and P9 peptide anchor preferences
in the B62 HLA supertype. The P2 peptide residue binds to the B-pocket in
HLA-B*1501. This pocket is relatively large because of the small Ser67 residue
located at the bottom. The peptide proximal part of the B-pocket is hydrophobic,
which is consistent with P2 anchor residue preference for Leu. The specificity
of the B-pocket is determined by the Met45, Ile66 and Ser67 residues. The apex
of the B-pocket is hydrophilic because of the Ser67 residue. The P9 peptide
residue binds to the F-pocket in HLA-B*1501. The residues most important for the
specificity of this pocket are Tyr74, Leu81, Leu95, Tyr123 and Trp147. These
residues create a hydrophobic interior in the F-pocket and their spatial
arrangement makes the pocket capable of containing large, bulky peptide side
chains. Ser116 is located at the bottom of the F-pocket and makes the bottom of
this pocket hydrophilic. Ser116, may act as a hydrogen-bonding partner and as
such is a perfect place for binding of a Tyr9 peptide residue. Thus, based on
structure information it is now possible to explain the peptide sequence
specificity of HLA-B*1501 as previously determined by peptide binding and pool
sequencing experiments.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 1.
Figure 1 ARP/wARP electron-density maps before manual inclusion
of the (a) LEKARGSTY and (b) ILGPPGSVY peptides from the
HLA-B*1501 complexes. The peptide N-termini are located at the
left and one-letter amino-acid codes are used. The
A-conformation of the ILGPPGSVY peptide is shown in red, while
the B-conformation is shown in green. The electron densities are
rendered at 1.0  level. level.
|
|
Figure 4.
Figure 4 Surface representations of (a) HLA-B*1501 (this work),
(b) HLA-A*0201 (Khan et al., 2000[Khan, A. R., Baker, B. M.,
Ghosh, P., Biddison, W. E. & Wiley, D. C. (2000). J. Immunol.
164, 6398-6405.]) and (c) HLA-A*1101 (Blicher et al.,
2005[Blicher, T., Kastrup, J. S., Buus, S. & Gajhede, M. (2005).
Acta Cryst. D61, 1031-1040.]). The surfaces are colored by the
standard convention for atoms.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from the IUCr:
Acta Crystallogr D Biol Crystallogr
(2006,
62,
1300-1310)
copyright 2006.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
P.T.Illing,
J.P.Vivian,
N.L.Dudek,
L.Kostenko,
Z.Chen,
M.Bharadwaj,
J.J.Miles,
L.Kjer-Nielsen,
S.Gras,
N.A.Williamson,
S.R.Burrows,
A.W.Purcell,
J.Rossjohn,
and
J.McCluskey
(2012).
Immune self-reactivity triggered by drug-modified HLA-peptide repertoire.
|
| |
Nature,
486,
554-558.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
A.Theodossis,
C.Guillonneau,
A.Welland,
L.K.Ely,
C.S.Clements,
N.A.Williamson,
A.I.Webb,
J.A.Wilce,
R.J.Mulder,
M.A.Dunstone,
P.C.Doherty,
J.McCluskey,
A.W.Purcell,
S.J.Turner,
and
J.Rossjohn
(2010).
Constraints within major histocompatibility complex class I restricted peptides: presentation and consequences for T-cell recognition.
|
| |
Proc Natl Acad Sci U S A,
107,
5534-5539.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
A.Wahl,
W.McCoy,
F.Schafer,
W.Bardet,
R.Buchli,
D.H.Fremont,
and
W.H.Hildebrand
(2009).
T-cell tolerance for variability in an HLA class I-presented influenza A virus epitope.
|
| |
J Virol,
83,
9206-9214.
|
|
|
|
|
|
|
G.Røder,
O.Kristensen,
J.S.Kastrup,
S.Buus,
and
M.Gajhede
(2008).
Structure of a SARS coronavirus-derived peptide bound to the human major histocompatibility complex class I molecule HLA-B*1501.
|
| |
Acta Crystallogr Sect F Struct Biol Cryst Commun,
64,
459-462.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
L.Sibilio,
A.Martayan,
A.Setini,
E.Lo Monaco,
E.Tremante,
R.H.Butler,
and
P.Giacomini
(2008).
A single bottleneck in HLA-C assembly.
|
| |
J Biol Chem,
283,
1267-1274.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
 |


 Links
Links















