 |
PDBsum entry 1vsu

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidoreductase
|
PDB id
|
|
|
|
1vsu
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.1.2.1.12
- glyceraldehyde-3-phosphate dehydrogenase (phosphorylating).
|
|
|
|
|
|
|
Pathway:
|
|
Glyceraldehyde-3-phosphate Dehydrogenase (phosphorylating)
|
|
|
|
|
|
Reaction:
|
|
D-glyceraldehyde 3-phosphate + phosphate + NAD+ = (2R)-3-phospho- glyceroyl phosphate + NADH + H+
|
|
|
|
|
|
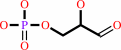 D-glyceraldehyde 3-phosphate
D-glyceraldehyde 3-phosphate
|
+
|
 phosphate
phosphate
|
+
|
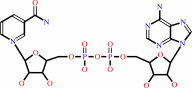 NAD(+)
NAD(+)
|
=
|
(2R)-3-phospho- glyceroyl phosphate
|
+
|
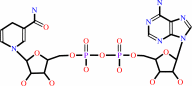 NADH
NADH
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
|
Bmc Struct Biol
9:9-9
(2009)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
An unexpected phosphate binding site in glyceraldehyde 3-phosphate dehydrogenase: crystal structures of apo, holo and ternary complex of Cryptosporidium parvum enzyme.
|
|
W.J.Cook,
O.Senkovich,
D.Chattopadhyay.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
BACKGROUND: The structure, function and reaction mechanism of glyceraldehyde
3-phosphate dehydrogenase (GAPDH) have been extensively studied. Based on these
studies, three anion binding sites have been identified, one 'Ps' site (for
binding the C-3 phosphate of the substrate) and two sites, 'Pi' and 'new Pi',
for inorganic phosphate. According to the original flip-flop model, the
substrate phosphate group switches from the 'Pi' to the 'Ps' site during the
multistep reaction. In light of the discovery of the 'new Pi' site, a modified
flip-flop mechanism, in which the C-3 phosphate of the substrate binds to the
'new Pi' site and flips to the 'Ps' site before the hydride transfer, was
proposed. An alternative model based on a number of structures of B.
stearothermophilus GAPDH ternary complexes (non-covalent and thioacyl
intermediate) proposes that in the ternary Michaelis complex the C-3 phosphate
binds to the 'Ps' site and flips from the 'Ps' to the 'new Pi' site during or
after the redox step. RESULTS: We determined the crystal structure of
Cryptosporidium parvum GAPDH in the apo and holo (enzyme + NAD) state and the
structure of the ternary enzyme-cofactor-substrate complex using an active site
mutant enzyme. The C. parvum GAPDH complex was prepared by pre-incubating the
enzyme with substrate and cofactor, thereby allowing free movement of the
protein structure and substrate molecules during their initial encounter.
Sulfate and phosphate ions were excluded from purification and crystallization
steps. The quality of the electron density map at 2A resolution allowed
unambiguous positioning of the substrate. In three subunits of the homotetramer
the C-3 phosphate group of the non-covalently bound substrate is in the 'new Pi'
site. A concomitant movement of the phosphate binding loop is observed in these
three subunits. In the fourth subunit the C-3 phosphate occupies an unexpected
site not seen before and the phosphate binding loop remains in the
substrate-free conformation. Orientation of the substrate with respect to the
active site histidine and serine (in the mutant enzyme) also varies in different
subunits. CONCLUSION: The structures of the C. parvum GAPDH ternary complex and
other GAPDH complexes demonstrate the plasticity of the substrate binding site.
We propose that the active site of GAPDH can accommodate the substrate in
multiple conformations at multiple locations during the initial encounter.
However, the C-3 phosphate group clearly prefers the 'new Pi' site for initial
binding in the active site.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|


 Links
Links











