 |
PDBsum entry 1v7z

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
PDB id:
|
|
|
|
| Name: |
|
Hydrolase
|
|
|
Title:
|
|
Creatininase-product complex
|
|
Structure:
|
|
Creatinine amidohydrolase. Chain: a, b, c, d, e, f. Synonym: creatininase. Engineered: yes
|
|
Source:
|
|
Pseudomonas sp.. Organism_taxid: 306. Expressed in: escherichia coli. Expression_system_taxid: 562.
|
|
Biol. unit:
|
|
Hexamer (from
)
|
|
Resolution:
|
|
|
1.60Å
|
R-factor:
|
0.183
|
R-free:
|
0.198
|
|
|
Authors:
|
|
T.Yoshimoto,N.Tanaka,N.Kanada,T.Inoue,Y.Nakajima,M.Haratake, K.T.Nakamura,Y.Xu,K.Ito
|
Key ref:

|
|
T.Yoshimoto
et al.
(2004).
Crystal structures of creatininase reveal the substrate binding site and provide an insight into the catalytic mechanism.
J Mol Biol,
337,
399-416.
PubMed id:
DOI:

|
|
|
Date:
|
|
|
26-Dec-03
|
Release date:
|
27-Jan-04
|
|
|
|
|
|
|
PROCHECK
|

|
|
|
|
|
Headers
|
|
|
|
References
|
|
|
|
|
|

|
|
P83772
(CRNA_PSEPU) -
Creatinine amidohydrolase from Pseudomonas putida
|
|
|
|
Seq:
Struc:
|
|
|
|
260 a.a.
257 a.a.
|
|

|

|
|
|
|
|
|
|
|
|
|
|
|
Key: |
 |
PfamA domain |
|
|
 |
Secondary structure |
|
 |
CATH domain |
|
|
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.3.5.2.10
- creatininase.
|
|
|
|
|
|
|
Reaction:
|
|
creatinine + H2O = creatine
|
|
|
|
|
|
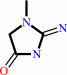 creatinine
creatinine
|
+
|
H2O
|
=
|
creatine
Bound ligand (Het Group name = )
corresponds exactly
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Mol Biol
337:399-416
(2004)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Crystal structures of creatininase reveal the substrate binding site and provide an insight into the catalytic mechanism.
|
|
T.Yoshimoto,
N.Tanaka,
N.Kanada,
T.Inoue,
Y.Nakajima,
M.Haratake,
K.T.Nakamura,
Y.Xu,
K.Ito.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Creatininase from Pseudomonas putida is a member of the urease-related
amidohydrolase superfamily. The crystal structure of the Mn-activated enzyme has
been solved by the single isomorphous replacement method at 1.8A resolution. The
structures of the native creatininase and the Mn-activated creatininase-creatine
complex have been determined by a difference Fourier method at 1.85 A and 1.6 A
resolution, respectively. We found the disc-shaped hexamer to be roughly 100 A
in diameter and 50 A in thickness and arranged as a trimer of dimers with 32
(D3) point group symmetry. The enzyme is a typical Zn2+ enzyme with a binuclear
metal center (metal1 and metal2). Atomic absorption spectrometry and X-ray
crystallography revealed that Zn2+ at metal1 (Zn1) was easily replaced with Mn2+
(Mn1). In the case of the Mn-activated enzyme, metal1 (Mn1) has a
square-pyramidal geometry bound to three protein ligands of Glu34, Asp45, and
His120 and two water molecules. Metal2 (Zn2) has a well-ordered tetrahedral
geometry bound to the three protein ligands of His36, Asp45, and Glu183 and a
water molecule. The crystal structure of the Mn-activated creatininase-creatine
complex, which is the first structure as the enzyme-substrate/inhibitor complex
of creatininase, reveals that significant conformation changes occur at the flap
(between the alpha5 helix and the alpha6 helix) of the active site and the
creatine is accommodated in a hydrophobic pocket consisting of Trp174, Trp154,
Tyr121, Phe182, Tyr153, and Gly119. The high-resolution crystal structure of the
creatininase-creatine complex enables us to identify two water molecules (Wat1
and Wat2) that are possibly essential for the catalytic mechanism of the enzyme.
The structure and proposed catalytic mechanism of the creatininase are different
from those of urease-related amidohydrolase superfamily enzymes. We propose a
new two-step catalytic mechanism possibly common to creatininases in which the
Wat1 acts as the attacking nucleophile in the water-adding step and the Wat2
acts as the catalytic acid in the ring-opening step.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 5.
Figure 5. Stereo diagrams showing the coordination of the
binuclear metal center of creatininase. The distances from the
metal ions to the ligands of the binuclear metal center of
creatininase and those from the water molecules to its ligands
are shown (in Å). (a) The Mn-activated creatininase. The
Mn1 has a square-pyramidal geometry, whereas the Zn2 is revealed
as having ordered tetrahedral geometry. (b) Native creatininase.
The Zn1 has distorted tetrahedral geometry, whereas the Zn2 has
ordered tetrahedral geometry.
|
|
Figure 9.
Figure 9. Proposed catalytic mechanism of creatininase.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Elsevier:
J Mol Biol
(2004,
337,
399-416)
copyright 2004.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
V.S.Lee,
K.Kodchakorn,
J.Jitonnom,
P.Nimmanpipug,
P.Kongtawelert,
and
B.Premanode
(2010).
Influence of metal cofactors and water on the catalytic mechanism of creatininase-creatinine in aqueous solution from molecular dynamics simulation and quantum study.
|
| |
J Comput Aided Mol Des,
24,
879-886.
|
|
|
|
|
|
|
T.Yoshimoto
(2007).
[Biochemistry and structural biology of microbial enzymes and their medical applications]
|
| |
Yakugaku Zasshi,
127,
1035-1045.
|
|
|
|
|
|
|
L.F.Huang,
B.Su,
S.C.Jao,
K.T.Liu,
and
W.S.Li
(2006).
Aminopeptidase p mediated detoxification of organophosphonate analogues of sarin: mechanistic and stereochemical study at the phosphorus atom of the substrate.
|
| |
Chembiochem,
7,
506-514.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
|
|




 Links
Links




