 |
PDBsum entry 1uxh

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidoreductase
|
PDB id
|
|
|
|
1uxh
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.1.1.1.37
- malate dehydrogenase.
|
|
|
|
|
|
|
Pathway:
|
|
Citric acid cycle
|
|
|
|
|
|
Reaction:
|
|
(S)-malate + NAD+ = oxaloacetate + NADH + H+
|
|
|
|
|
|
(S)-malate
Bound ligand (Het Group name = )
matches with 88.89% similarity
|
+
|
NAD(+)
Bound ligand (Het Group name = )
corresponds exactly
|
=
|
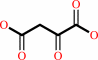 oxaloacetate
oxaloacetate
|
+
|
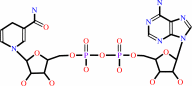 NADH
NADH
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Mol Biol
341:1215-1226
(2004)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Large improvement in the thermal stability of a tetrameric malate dehydrogenase by single point mutations at the dimer-dimer interface.
|
|
A.Bjørk,
B.Dalhus,
D.Mantzilas,
R.Sirevåg,
V.G.Eijsink.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The stability of tetrameric malate dehydrogenase from the green phototrophic
bacterium Chloroflexus aurantiacus (CaMDH) is at least in part determined by
electrostatic interactions at the dimer-dimer interface. Since previous studies
had indicated that the thermal stability of CaMDH becomes lower with increasing
pH, attempts were made to increase the stability by removal of (excess) negative
charge at the dimer-dimer interface. Mutation of Glu165 to Gln or Lys yielded a
dramatic increase in thermal stability at pH 7.5 (+23.6 -- + 23.9 degrees C
increase in apparent t(m)) and a more moderate increase at pH 4.4 (+4.6 -- + 5.4
degrees C). The drastically increased stability at neutral pH was achieved
without forfeiture of catalytic performance at low temperatures. The crystal
structures of the two mutants showed only minor structural changes close to the
mutated residues, and indicated that the observed stability effects are solely
due to subtle changes in the complex network of electrostatic interactions in
the dimer-dimer interface. Both mutations reduced the concentration dependency
of thermal stability, suggesting that the oligomeric structure had been
reinforced. Interestingly, the two mutations had similar effects on stability,
despite the charge difference between the introduced side-chains. Together with
the loss of concentration dependency, this may indicate that both E165Q and
E165K stabilize CaMDH to such an extent that disruption of the inter-dimer
electrostatic network around residue 165 no longer limits kinetic thermal
stability.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 1.
Figure 1. The structure of homotetrameric MDH from C. aurantiacus is shown in a ribbon diagram in the upper panel.
The tetramer consists of two dimers (AB and CD). The NAD cofactor bound to the catalytic centre is shown in yellow.
The rectangle indicates the location of the electrostatic network around Glu165 at the dimer--dimer interface in CaMDH
(note that the network occurs four times in the tetramer). The lower panel shows a stereo view of the environment of
Glu165 in subunit A, with backbones coloured according to the scheme used in the upper panel. The side-chains are
shown in ball and stick in atom colours, Cd
2C
bound to Glu165 is shown in yellow. Dotted lines indicate interacting
atoms, located within 3 Å of each other.
|
|
Figure 5.
Figure 5. Stereo pictures showing the environment of residue 165 in cadmium-containing structures of wild-type
CaMDH (A) and the E165Q (B) and E165K (C) mutants. (B) and (C) show 2F oKFc maps contoured at 1s for residue 165 to
illustrate the conformation of the side-chains as well as the general quality of the refinement. Red spheres represent
water molecules; the yellow sphere represents cadmium. In CaMDH (A) the negatively charged Glu165 binds directly to
one of the cadmium ions in the structure (closest distance 2.5 A
�
). In the E165Q mutant (B) the cadmium ion binds to
residue 165 via a water molecule (closest Cd
2C
-165 distance 4.2 A
�
). In the E165K mutant (A) the cadmium ion has moved
even further away from its original position and is now very close (2.3 A
�
, as compared to 4.2 A
�
in the wild-type) to
Glu238 of the other monomer. The distance between cadmium and the z-amino group of Lys165 is 4.1 A
�
. Figure 1 and
this Figure were made using the programs Bobscript
64
and Raster 3D.
65
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Elsevier:
J Mol Biol
(2004,
341,
1215-1226)
copyright 2004.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
G.A.Kotzia,
and
N.E.Labrou
(2009).
Engineering thermal stability of L-asparaginase by in vitro directed evolution.
|
| |
FEBS J,
276,
1750-1761.
|
|
|
|
|
|
|
Z.Xiao,
H.Bergeron,
S.Grosse,
M.Beauchemin,
M.L.Garron,
D.Shaya,
T.Sulea,
M.Cygler,
and
P.C.Lau
(2008).
Improvement of the thermostability and activity of a pectate lyase by single amino acid substitutions, using a strategy based on melting-temperature-guided sequence alignment.
|
| |
Appl Environ Microbiol,
74,
1183-1189.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
A.F.Mehl,
B.Demeler,
and
A.Zraikat
(2007).
A water mediated electrostatic interaction gives thermal stability to the "tail" region of the GrpE protein from E. coli.
|
| |
Protein J,
26,
239-245.
|
|
|
|
|
|
|
N.Zheng,
B.Huang,
J.Xu,
S.Huang,
J.Chen,
X.Hu,
K.Ying,
and
X.Yu
(2006).
Enzymatic and physico-chemical characteristics of recombinant cMDH and mMDH of Clonorchis sinensis.
|
| |
Parasitol Res,
99,
174-180.
|
|
|
|
|
|
|
V.G.Eijsink,
S.Gåseidnes,
T.V.Borchert,
and
B.van den Burg
(2005).
Directed evolution of enzyme stability.
|
| |
Biomol Eng,
22,
21-30.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|


 Links
Links













