 |
PDBsum entry 1t4z

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.2.7.13.3
- histidine kinase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + protein L-histidine = ADP + protein N-phospho-L-histidine
|
|
|
|
|
|
 ATP
ATP
|
+
|
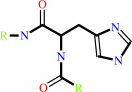 protein L-histidine
protein L-histidine
|
=
|
 ADP
ADP
|
+
|
protein N-phospho-L-histidine
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Mol Biol
342:9
(2004)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Structure of the N-terminal domain of the circadian clock-associated histidine kinase SasA.
|
|
I.Vakonakis,
D.A.Klewer,
S.B.Williams,
S.S.Golden,
A.C.LiWang.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Circadian oscillators are endogenous biological systems that generate the
approximately 24 hour temporal pattern of biological processes and confer a
reproductive fitness advantage to their hosts. The cyanobacterial clock is the
simplest known and the only clock system for which structural information for
core component proteins, in this case KaiA, KaiB and KaiC, is available. SasA, a
clock-associated histidine kinase, is necessary for robustness of the circadian
rhythm of gene expression and implicated in clock output. The N-terminal domain
of SasA (N-SasA) interacts directly with KaiC and likely functions as the
sensory domain controlling the SasA histidine kinase activity. N-SasA and KaiB
share significant sequence similarity and, thus, it has been proposed that they
would be structurally similar and may even compete for KaiC binding. Here, we
report the NMR structure of N-SasA and show it to be different from that of
KaiB. The structural comparisons provide no clear details to suggest competition
of SasA and KaiB for KaiC binding. N-SasA adopts a canonical thioredoxin fold
but lacks the catalytic cysteine residues. A patch of conserved, solvent-exposed
residues is found near the canonical thioredoxin active site. We suggest that
this surface is used by N-SasA for protein-protein interactions. Our analysis
suggests that the structural differences between N-SasA and KaiB are the result
of only a few critical amino acid substitutions.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 2.
Figure 2. Solution structure of the N-SasA domain and
comparison to other thioredoxin-fold proteins. Shown here are
(A) the average minimized structure of N-SasA and (B) the
25-structure ensemble in two mutually orthogonal views. The N-
and C-termini of the protein are indicated. The ensemble of
structures was superimposed using the backbone of low-mobility
residues (Table 1). (C) Comparison of bacteriophage T4
glutaredoxin (PDB-ID 1AAZ), the N-SasA domain and Escherichia
coli thioredoxin (2TRX). The unstructured first ten residues of
N-SasA, and the first 20 residues of thioredoxin that have no
structural homologues in N-SasA are not shown. The structures
were superimposed using the C^a atoms of the b-sheet.
|
|
Figure 4.
Figure 4. Structure comparison of the N-SasA domain and
KaiB. (A) Shown here is a CLUSTAL-W alignment of S. elongatus
N-SasA and KaiB. The secondary structure elements from the
solution structure of N-SasA (top line), and the crystal
structure of the Anabaena sp. PCC 7120 KaiB8 (highly homologous
to S. elongatus KaiB; bottom line) are shown. Residues likely to
be critical for the differences in the two structures are
indicated by yellow bars. (B) Secondary structure prediction for
S. elongatus N-SasA and KaiB. Prediction was performed by
PSIPRED36 and the regular secondary structure elements
identified are shown. N-SasA residues 1-8 are not shown. (C)
Structures and (D) protein architectures of S. elongatus N-SasA
and Anabaena sp. KaiB.8 The unstructured first ten residues of
N-SasA are not shown. The protein architectures are highly
similar for the first half of the sequences but diverge in the
second half.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Elsevier:
J Mol Biol
(2004,
342,
9-0)
copyright 2004.
|
|
| |
Figures were
selected
by the author.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
T.L.Wood,
J.Bridwell-Rabb,
Y.I.Kim,
T.Gao,
Y.G.Chang,
A.LiWang,
D.P.Barondeau,
and
S.S.Golden
(2010).
The KaiA protein of the cyanobacterial circadian oscillator is modulated by a redox-active cofactor.
|
| |
Proc Natl Acad Sci U S A,
107,
5804-5809.
|
|
|
|
|
|
|
S.S.Golden
(2007).
Integrating the circadian oscillator into the life of the cyanobacterial cell.
|
| |
Cold Spring Harb Symp Quant Biol,
72,
331-338.
|
|
|
|
|
|
|
T.Gao,
X.Zhang,
N.B.Ivleva,
S.S.Golden,
and
A.LiWang
(2007).
NMR structure of the pseudo-receiver domain of CikA.
|
| |
Protein Sci,
16,
465-475.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
A.Andreeva,
and
A.G.Murzin
(2006).
Evolution of protein fold in the presence of functional constraints.
|
| |
Curr Opin Struct Biol,
16,
399-408.
|
|
|
|
|
|
|
M.Nouailler,
X.Morelli,
O.Bornet,
B.Chetrit,
Z.Dermoun,
and
F.Guerlesquin
(2006).
Solution structure of HndAc: a thioredoxin-like domain involved in the NADP-reducing hydrogenase complex.
|
| |
Protein Sci,
15,
1369-1378.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
R.M.Smith,
and
S.B.Williams
(2006).
Circadian rhythms in gene transcription imparted by chromosome compaction in the cyanobacterium Synechococcus elongatus.
|
| |
Proc Natl Acad Sci U S A,
103,
8564-8569.
|
|
|
|
|
|
|
K.Hitomi,
T.Oyama,
S.Han,
A.S.Arvai,
and
E.D.Getzoff
(2005).
Tetrameric architecture of the circadian clock protein KaiB. A novel interface for intermolecular interactions and its impact on the circadian rhythm.
|
| |
J Biol Chem,
280,
19127-19135.
|
|
|
PDB code:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
code is
shown on the right.
|
|


 Links
Links











