 |
PDBsum entry 1n0h

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.2.2.1.6
- acetolactate synthase.
|
|
|
|
|
|
|
Pathway:
|
|
Isoleucine and Valine Biosynthesis
|
|
|
|
|
|
Reaction:
|
|
2 pyruvate + H+ = (2S)-2-acetolactate + CO2
|
|
|
|
|
|
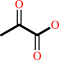 2
×
pyruvate
2
×
pyruvate
|
+
|
H(+)
Bound ligand (Het Group name = )
matches with 55.56% similarity
|
=
|
(2S)-2-acetolactate
|
+
|
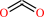 CO2
CO2
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
Thiamine diphosphate
|
|
|
|
|
|
Thiamine diphosphate
Bound ligand (Het Group name =
TPP)
corresponds exactly
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Biol Chem
279:2242-2253
(2004)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
The crystal structures of Klebsiella pneumoniae acetolactate synthase with enzyme-bound cofactor and with an unusual intermediate.
|
|
S.S.Pang,
R.G.Duggleby,
R.L.Schowen,
L.W.Guddat.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Acetohydroxyacid synthase (AHAS) and acetolactate synthase (ALS) are thiamine
diphosphate (ThDP)-dependent enzymes that catalyze the decarboxylation of
pyruvate to give a cofactor-bound hydroxyethyl group, which is transferred to a
second molecule of pyruvate to give 2-acetolactate. AHAS is found in plants,
fungi, and bacteria, is involved in the biosynthesis of the branched-chain amino
acids, and contains non-catalytic FAD. ALS is found only in some bacteria, is a
catabolic enzyme required for the butanediol fermentation, and does not contain
FAD. Here we report the 2.3-A crystal structure of Klebsiella pneumoniae ALS.
The overall structure is similar to AHAS except for a groove that accommodates
FAD in AHAS, which is filled with amino acid side chains in ALS. The ThDP
cofactor has an unusual conformation that is unprecedented among the 26 known
three-dimensional structures of nine ThDP-dependent enzymes, including AHAS.
This conformation suggests a novel mechanism for ALS. A second structure, at 2.0
A, is described in which the enzyme is trapped halfway through the catalytic
cycle so that it contains the hydroxyethyl intermediate bound to ThDP. The
cofactor has a tricyclic structure that has not been observed previously in any
ThDP-dependent enzyme, although similar structures are well known for free
thiamine. This structure is consistent with our proposed mechanism and probably
results from an intramolecular proton transfer within a tricyclic carbanion that
is the true reaction intermediate. Modeling of the second molecule of pyruvate
into the active site of the enzyme with the bound intermediate is consistent
with the stereochemistry and specificity of ALS.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 2.
FIG. 2. Structure of K. pneumoniae ALS. A shows a ribbon
diagram of the overall structure of the resting enzyme tetramer,
with the monomers colored green (monomer A), red (B), blue (C),
and yellow (D). There is a vertical 2-fold axis of symmetry in
this view. The asymmetric unit contains monomers A and B,
whereas the active sites are at the AC and BD interfaces.
Monomer A is shown in B, with cylinder representations of  -helices
(red) and -helices
(red) and  -strands (turquoise)
shown as arrows, connected by random coil (green). C and D
compare the -domains of ALS (C) and
AHAS (D), with secondary structure indicated as in B. The
residues in contact with FAD (stick model, D) in AHAS and their
structural equivalents in ALS (C) are shown in surface
representation. -strands (turquoise)
shown as arrows, connected by random coil (green). C and D
compare the -domains of ALS (C) and
AHAS (D), with secondary structure indicated as in B. The
residues in contact with FAD (stick model, D) in AHAS and their
structural equivalents in ALS (C) are shown in surface
representation.
|
|
Figure 7.
FIG. 7. The active sites of ALS and AHAS. A shows the
active site region of K. pneumoniae ALS (resting enzyme), and B
shows the active site of yeast AHAS viewed in a similar
orientation. Residues with and without the prime symbol are
derived from different monomers. C, the active site of ALS is
shown with the second molecule of pyruvate modeled in so that it
makes favorable contacts and is orientated so that it would
yield the S-enantiomer of acetolactate (D). In this model, the
intermediate is represented as the tricyclic carbanion (IVb,
Fig. 5), and an alternate conformation of Lys36 is shown for
which the  -amino group forms an
ionic interaction with the carboxylate of the second pyruvate. -amino group forms an
ionic interaction with the carboxylate of the second pyruvate.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from the ASBMB:
J Biol Chem
(2004,
279,
2242-2253)
copyright 2004.
|
|
| |
Figures were
selected
by the author.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
I.Hartmann,
P.Carranza,
A.Lehner,
R.Stephan,
L.Eberl,
and
K.Riedel
(2010).
Genes involved in Cronobacter sakazakii biofilm formation.
|
| |
Appl Environ Microbiol,
76,
2251-2261.
|
|
|
|
|
|
|
S.Atsumi,
Z.Li,
and
J.C.Liao
(2009).
Acetolactate synthase from Bacillus subtilis serves as a 2-ketoisovalerate decarboxylase for isobutanol biosynthesis in Escherichia coli.
|
| |
Appl Environ Microbiol,
75,
6306-6311.
|
|
|
|
|
|
|
E.P.Balskus,
and
C.T.Walsh
(2008).
Investigating the initial steps in the biosynthesis of cyanobacterial sunscreen scytonemin.
|
| |
J Am Chem Soc,
130,
15260-15261.
|
|
|
|
|
|
|
W.Gong,
B.Hao,
Z.Wei,
D.J.Ferguson,
T.Tallant,
J.A.Krzycki,
and
M.K.Chan
(2008).
Structure of the alpha2epsilon2 Ni-dependent CO dehydrogenase component of the Methanosarcina barkeri acetyl-CoA decarbonylase/synthase complex.
|
| |
Proc Natl Acad Sci U S A,
105,
9558-9563.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
E.C.Juan,
M.M.Hoque,
M.T.Hossain,
T.Yamamoto,
S.Imamura,
K.Suzuki,
T.Sekiguchi,
and
A.Takénaka
(2007).
The structures of pyruvate oxidase from Aerococcus viridans with cofactors and with a reaction intermediate reveal the flexibility of the active-site tunnel for catalysis.
|
| |
Acta Crystallogr Sect F Struct Biol Cryst Commun,
63,
900-907.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
T.Ohsako,
and
T.Tominaga
(2007).
Nucleotide substitutions in the acetolactate synthase genes of sulfonylurea-resistant biotypes of Monochoria vaginalis (Pontederiaceae).
|
| |
Genes Genet Syst,
82,
207-215.
|
|
|
|
|
|
|
J.A.McCourt,
and
R.G.Duggleby
(2006).
Acetohydroxyacid synthase and its role in the biosynthetic pathway for branched-chain amino acids.
|
| |
Amino Acids,
31,
173-210.
|
|
|
|
|
|
|
D.M.Chipman,
R.G.Duggleby,
and
K.Tittmann
(2005).
Mechanisms of acetohydroxyacid synthases.
|
| |
Curr Opin Chem Biol,
9,
475-481.
|
|
|
|
|
|
|
R.Paulini,
K.Müller,
and
F.Diederich
(2005).
Orthogonal multipolar interactions in structural chemistry and biology.
|
| |
Angew Chem Int Ed Engl,
44,
1788-1805.
|
|
|
|
|
|
|
T.G.Mosbacher,
M.Mueller,
and
G.E.Schulz
(2005).
Structure and mechanism of the ThDP-dependent benzaldehyde lyase from Pseudomonas fluorescens.
|
| |
FEBS J,
272,
6067-6076.
|
|
|
PDB codes:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
code is
shown on the right.
|
|




 Links
Links













