 |
PDBsum entry 1dso

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidoreductase
|
PDB id
|
|
|
|
1dso
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.1.11.1.5
- cytochrome-c peroxidase.
|
|
|
|
|
|
|
Reaction:
|
|
2 Fe(II)-[cytochrome c] + H2O2 + 2 H+ = 2 Fe(III)-[cytochrome c] + 2 H2O
|
|
|
|
|
|
2
×
Fe(II)-[cytochrome c]
|
+
|
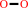 H2O2
H2O2
|
+
|
2
×
H(+)
|
=
|
2
×
Fe(III)-[cytochrome c]
|
+
|
2
×
H2O
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
Heme
|
|
|
|
|
|
Heme
Bound ligand (Het Group name =
HEM)
matches with 95.45% similarity
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Biochemistry
40:1265-1273
(2001)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Replacement of the axial histidine ligand with imidazole in cytochrome c peroxidase. 1. Effects on structure.
|
|
J.Hirst,
S.K.Wilcox,
P.A.Williams,
J.Blankenship,
D.E.McRee,
D.B.Goodin.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Replacement of the axial histidine ligand with exogenous imidazole has been
accomplished in a number of heme protein mutants, where it often serves to
complement the functional properties of the protein. In this paper, we describe
the effects of pH and buffer ion on the crystal structure of the H175G mutant of
cytochrome c peroxidase, in which the histidine tether between the heme and the
protein backbone is replaced by bound imidazole. The structures show that
imidazole can occupy the proximal H175G cavity under a number of experimental
conditions, but that the details of the interaction with the protein and the
coordination to the heme are markedly dependent on conditions. Replacement of
the tethered histidine ligand with imidazole permits the heme to shift slightly
in its pocket, allowing it to adopt either a planar or distally domed
conformation. H175G crystallized from both high phosphate and imidazole
concentrations exists as a novel, 5-coordinate phosphate bound state, in which
the proximal imidazole is dissociated and the distal phosphate is coordinated to
the iron. To accommodate this bound phosphate, the side chains of His-52 and
Asn-82 alter their positions and a significant conformational change in the
surrounding protein backbone occurs. In the absence of phosphate, imidazole
binds to the proximal H175G cavity in a pH-dependent fashion. At pH 7, imidazole
is directly coordinated to the heme (d(Fe--Im) = 2.0 A) with a nearby distal
water (d(Fe--HOH) = 2.4 A). This is similar to the structure of WT CCP except
that the iron lies closer in the heme plane, and the hydrogen bond between
imidazole and Asp-235 (d(Im--Asp) = 3.1 A) is longer than for WT CCP
(d(His--Asp) = 2.9 A). As the pH is dropped to 5, imidazole dissociates from the
heme (d(Fe--Im) = 2.9 A), but remains in the proximal cavity where it is
strongly hydrogen bonded to Asp-235 (d(Im--Asp) = 2.8 A). In addition, the heme
is significantly domed toward the distal pocket where it may coordinate a water
molecule. Finally, the structure of H175G/Im, pH 6, at low temperature (100 K)
is very similar to that at room temperature, except that the water above the
distal heme face is not present. This study concludes that steric restrictions
imposed by the covalently tethered histidine restrain the heme and its ligand
coordination from distortions that would arise in the absence of the restricted
tether. Coupled with the functional and spectroscopic properties described in
the following paper in this issue, these structures help to illustrate how the
delicate and critical interactions between protein, ligand, and metal modulate
the function of heme enzymes.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
S.W.Vetter,
A.C.Terentis,
R.L.Osborne,
J.H.Dawson,
and
D.B.Goodin
(2009).
Replacement of the axial histidine heme ligand with cysteine in nitrophorin 1: spectroscopic and crystallographic characterization.
|
| |
J Biol Inorg Chem,
14,
179-191.
|
|
|
|
|
|
|
K.Kobayashi,
M.Kubo,
S.Yoshioka,
T.Kitagawa,
Y.Kato,
Y.Asano,
and
S.Aono
(2006).
Systematic regulation of the enzymatic activity of phenylacetaldoxime dehydratase by exogenous ligands.
|
| |
Chembiochem,
7,
2004-2009.
|
|
|
|
|
|
|
A.M.Hays,
H.B.Gray,
and
D.B.Goodin
(2003).
Trapping of peptide-based surrogates in an artificially created channel of cytochrome c peroxidase.
|
| |
Protein Sci,
12,
278-287.
|
|
|
|
|
|
|
R.J.Rosenfeld,
A.M.Hays,
R.A.Musah,
and
D.B.Goodin
(2002).
Excision of a proposed electron transfer pathway in cytochrome c peroxidase and its replacement by a ligand-binding channel.
|
| |
Protein Sci,
11,
1251-1259.
|
|
|
PDB codes:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|


 Links
Links











