 |
PDBsum entry 1bwk

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidoreductase
|
PDB id
|
|
|
|
1bwk
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.1.6.99.1
- Nadph dehydrogenase.
|
|
|
|
|
|
|
Reaction:
|
|
A + NADPH + H+ = AH2 + NADP+
|
|
|
|
|
|
|
+
|
 NADPH
NADPH
|
+
|
H(+)
|
=
|
AH2
|
+
|
 NADP(+)
NADP(+)
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
FMN or FAD
|
|
|
|
|
|
FMN
Bound ligand (Het Group name =
FMN)
corresponds exactly
|
or
|
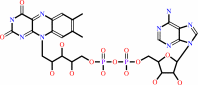 FAD
FAD
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Biol Chem
273:32753-32762
(1998)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
On the active site of Old Yellow Enzyme. Role of histidine 191 and asparagine 194.
|
|
B.J.Brown,
Z.Deng,
P.A.Karplus,
V.Massey.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Old Yellow Enzyme (OYE) binds phenolic ligands forming long wavelength (500-800
nm) charge-transfer complexes. The enzyme is reduced by NADPH, and oxygen,
quinones, and alpha,beta-unsaturated aldehydes and ketones can act as electron
acceptors to complete catalytic turnover. Solution of the crystal structure of
OYE1 from brewer's bottom yeast (Fox, K. M., and Karplus, P. A. (1994) Structure
2, 1089-1105) made it possible to identify histidine 191 and asparagine 194 as
amino acid residues that hydrogen-bond with the phenolic ligands, stabilizing
the anionic form involved in charge-transfer interaction with the FMN prosthetic
group. His-191 and Asn-194 are also predicted to interact with the nicotinamide
ring of NADPH in the active site. Mutations of His-191 to Asn, Asn-194 to His,
and a double mutation, H191N/N194H, were made of OYE1. It was not possible to
isolate the N191H mutant enzyme, but the other two mutant forms had the expected
effect on phenolic ligand binding, i.e. decreased binding affinity and decreased
charge-transfer absorbance. Reduction of the H191N mutant enzyme by NADPH was
similar to that of OYE1, but the reduction rate constant for NADH was greatly
decreased. The double mutant enzyme had an increased rate constant for reduction
by NADPH, but the reduction rate constant with NADH was lower by a factor of 15.
The reactivity of OYE1 and the mutant enzymes with oxygen was similar, but the
reactivity of 2-cyclohexenone was greatly decreased by the mutations. The
crystal structures of the two mutant forms showed only minor changes from that
of the wild type enzyme.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 5.
Fig. 5. Oxidation of OYE1 by 2-cyclohexenone. OYE1 was
reduced by an NADPH-generating system consisting of
glucose-6-phosphate (0.625 mM), glucose-6-phosphate
dehydrogenase (5 µl), and NADP (0.625 µM). Reduction
of the enzyme was followed spectrally and was complete at 84
min. The reduced enzyme was mixed in the stopped flow apparatus
with the concentrations of cyclohexenone shown, and the reaction
traces at 460 nm were used to calculate the values of k[obs]
shown.
|
|
Figure 6.
Fig. 6. Active site of OYE1. Overlay of four structures
around the active sites: PHB bound wild type OYE1 as the
thickest lines (PDB entry code 1OYB), empty wild type structure
(PDB entry code 1OYA) as medium thickness lines, H191N as dashed
lines, and H191N/N194H as thin lines.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from the ASBMB:
J Biol Chem
(1998,
273,
32753-32762)
copyright 1998.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
H.S.Toogood,
A.Fryszkowska,
M.Hulley,
M.Sakuma,
D.Mansell,
G.M.Stephens,
J.M.Gardiner,
and
N.S.Scrutton
(2011).
A site-saturated mutagenesis study of pentaerythritol tetranitrate reductase reveals that residues 181 and 184 influence ligand binding, stereochemistry and reactivity.
|
| |
Chembiochem,
12,
738-749.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
R.Pandey,
S.Ghosh,
S.Mukhopadhyay,
S.Ramasesha,
and
P.K.Das
(2011).
Geometry and quadratic nonlinearity of charge transfer complexes in solution using depolarized hyper-Rayleigh scattering.
|
| |
J Chem Phys,
134,
044533.
|
|
|
|
|
|
|
W.Li,
F.Zhou,
B.Liu,
D.Feng,
Y.He,
K.Qi,
H.Wang,
and
J.Wang
(2011).
Comparative characterization, expression pattern and function analysis of the 12-oxo-phytodienoic acid reductase gene family in rice.
|
| |
Plant Cell Rep,
30,
981-995.
|
|
|
|
|
|
|
E.M.Koehn,
and
A.Kohen
(2010).
Flavin-dependent thymidylate synthase: a novel pathway towards thymine.
|
| |
Arch Biochem Biophys,
493,
96.
|
|
|
|
|
|
|
E.M.Koehn,
T.Fleischmann,
J.A.Conrad,
B.A.Palfey,
S.A.Lesley,
I.I.Mathews,
and
A.Kohen
(2009).
An unusual mechanism of thymidylate biosynthesis in organisms containing the thyX gene.
|
| |
Nature,
458,
919-923.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
D.J.Kang,
J.M.Ridlon,
D.R.Moore,
S.Barnes,
and
P.B.Hylemon
(2008).
Clostridium scindens baiCD and baiH genes encode stereo-specific 7alpha/7beta-hydroxy-3-oxo-delta4-cholenoic acid oxidoreductases.
|
| |
Biochim Biophys Acta,
1781,
16-25.
|
|
|
|
|
|
|
P.van Dillewijn,
R.M.Wittich,
A.Caballero,
and
J.L.Ramos
(2008).
Subfunctionality of hydride transferases of the old yellow enzyme family of flavoproteins of Pseudomonas putida.
|
| |
Appl Environ Microbiol,
74,
6703-6708.
|
|
|
|
|
|
|
A.Müller,
B.Hauer,
and
B.Rosche
(2007).
Asymmetric alkene reduction by yeast old yellow enzymes and by a novel Zymomonas mobilis reductase.
|
| |
Biotechnol Bioeng,
98,
22-29.
|
|
|
|
|
|
|
C.Breithaupt,
R.Kurzbauer,
H.Lilie,
A.Schaller,
J.Strassner,
R.Huber,
P.Macheroux,
and
T.Clausen
(2006).
Crystal structure of 12-oxophytodienoate reductase 3 from tomato: self-inhibition by dimerization.
|
| |
Proc Natl Acad Sci U S A,
103,
14337-14342.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
H.Khan,
T.Barna,
N.C.Bruce,
A.W.Munro,
D.Leys,
and
N.S.Scrutton
(2005).
Proton transfer in the oxidative half-reaction of pentaerythritol tetranitrate reductase. Structure of the reduced enzyme-progesterone complex and the roles of residues Tyr186, His181, His184.
|
| |
FEBS J,
272,
4660-4671.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
S.J.Marshall,
D.Krause,
D.K.Blencowe,
and
G.F.White
(2004).
Characterization of glycerol trinitrate reductase (NerA) and the catalytic role of active-site residues.
|
| |
J Bacteriol,
186,
1802-1810.
|
|
|
|
|
|
|
C.Breithaupt,
J.Strassner,
U.Breitinger,
R.Huber,
P.Macheroux,
A.Schaller,
and
T.Clausen
(2001).
X-ray structure of 12-oxophytodienoate reductase 1 provides structural insight into substrate binding and specificity within the family of OYE.
|
| |
Structure,
9,
419-429.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
Y.Meah,
B.J.Brown,
S.Chakraborty,
and
V.Massey
(2001).
Old yellow enzyme: reduction of nitrate esters, glycerin trinitrate, and propylene 1,2-dinitrate.
|
| |
Proc Natl Acad Sci U S A,
98,
8560-8565.
|
|
|
|
|
|
|
J.Buckman,
and
S.M.Miller
(2000).
Transient kinetics and intermediates formed during the electron transfer reaction catalyzed by Candida albicans estrogen binding protein.
|
| |
Biochemistry,
39,
10521-10531.
|
|
|
|
|
|
|
Y.Meah,
and
V.Massey
(2000).
Old yellow enzyme: stepwise reduction of nitro-olefins and catalysis of aci-nitro tautomerization.
|
| |
Proc Natl Acad Sci U S A,
97,
10733-10738.
|
|
|
|
|
|
|
D.Xu,
R.M.Kohli,
and
V.Massey
(1999).
The role of threonine 37 in flavin reactivity of the old yellow enzyme.
|
| |
Proc Natl Acad Sci U S A,
96,
3556-3561.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|


 Links
Links













