 |
PDBsum entry 3fbx
|
|
|
|
References listed in PDB file
|
 |
|
Key reference
|
|
|
Title
|
|
De novo sulfur sad phasing of the lysosomal 66.3 kda protein from mouse.
|
|
|
Authors
|
|
K.Lakomek,
A.Dickmanns,
U.Mueller,
K.Kollmann,
F.Deuschl,
A.Berndt,
T.Lübke,
R.Ficner.
|
|
|
Ref.
|
|
Acta Crystallogr D Biol Crystallogr, 2009,
65,
220-228.
[DOI no: ]
|
|
|
PubMed id
|
|
|
|
|
|
Abstract
|
|
|
The 66.3 kDa protein from mouse is a soluble protein of the lysosomal matrix. It
is synthesized as a glycosylated 75 kDa preproprotein which is further processed
into 28 and 40 kDa fragments. Despite bioinformatics approaches and molecular
characterization of the 66.3 kDa protein, the mode of its maturation as well as
its physiological function remained unknown. Therefore, it was decided to tackle
this question by means of X-ray crystallography. After expression in a human
fibrosarcoma cell line, the C-terminally His-tagged single-chain 66.3 kDa
variant and the double-chain form consisting of a 28 kDa fragment and a 40 kDa
fragment were purified to homogeneity but could not be separated during the
purification procedure. This mixture was therefore used for crystallization.
Single crystals were obtained and the structure of the 66.3 kDa protein was
solved by means of sulfur SAD phasing using data collected at a wavelength of
1.9 A on the BESSY beamline BL14.2 of Freie Universität Berlin. Based on the
anomalous signal, a 22-atom substructure comprising 21 intrinsic S atoms and one
Xe atom with very low occupancy was found and refined at a resolution of 2.4 A
using the programs SHELXC/D and SHARP. Density modification using SOLOMON and DM
resulted in a high-quality electron-density map, enabling automatic model
building with ARP/wARP. The initial model contained 85% of the amino-acid
residues expected to be present in the asymmetric unit of the crystal.
Subsequently, the model was completed and refined to an R(free) factor of 19.8%.
The contribution of the single Xe atom to the anomalous signal was analyzed in
comparison to that of the S atoms and was found to be negligible. This work
should encourage the use of the weak anomalous scattering of intrinsic S atoms
in SAD phasing, especially for proteins, which require both expensive and
time-consuming expression and purification procedures, preventing extensive
screening of heavy-atom crystal soaks.
|
|
|
|

|
|

|
|
Figure 3.
Figure 3 Anomalous difference Patterson map. Sharpened map of
the Harker section v = 0 calculated at a resolution of 2.4
Å. Contours are in increments of 1.3  and
are coloured differently for each level. and
are coloured differently for each level.
|
|
Figure 4.
Figure 4 Comparison of the experimental electron-density maps
after density modification providing SHARP with input parameters
with heavy-atom sites including and lacking the Xe atom,
respectively. The maps are coloured blue and green,
respectively, and are contoured at the 1.5 level.
(a) The section encompassing several neighbouring molecules
shows a clear solvent boundary. (b) The 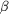 -strands
forming a -sheet
are well defined in the electron-density map. (c, d) Density for
an -strands
forming a -sheet
are well defined in the electron-density map. (c, d) Density for
an  -helical
structure is also visible, viewed from the top (c) and from the
side (d) of the helix, respectively. (e, f) Close-up views of
the side chains of Lys340 at the surface (e) and Phe371 in the
hydrophobic core of the protein (f), respectively. -helical
structure is also visible, viewed from the top (c) and from the
side (d) of the helix, respectively. (e, f) Close-up views of
the side chains of Lys340 at the surface (e) and Phe371 in the
hydrophobic core of the protein (f), respectively.
|
|
|
|
|
|
The above figures are
reprinted
by permission from the IUCr:
Acta Crystallogr D Biol Crystallogr
(2009,
65,
220-228)
copyright 2009.
|
|
|
|
|
|
|




