 |
PDBsum entry 2a5i

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
PDB id:
|
|
|
|
| Name: |
|
Hydrolase
|
|
|
Title:
|
|
Crystal structures of sars coronavirus main peptidase inhibited by an aza-peptide epoxide in the space group c2
|
|
Structure:
|
|
3c-like peptidase. Chain: a. Synonym: 3cl-pro, 3clp, nsp2. Engineered: yes
|
|
Source:
|
|
Sars coronavirus. Organism_taxid: 227859. Strain: sars. Expressed in: escherichia coli. Expression_system_taxid: 562.
|
|
Resolution:
|
|
|
1.88Å
|
R-factor:
|
0.200
|
R-free:
|
0.242
|
|
|
Authors:
|
|
T.-W.Lee,M.M.Cherney,C.Huitema,J.Liu,K.E.James,J.C.Powers,L.D.Eltis, M.N.James
|
Key ref:

|
|
T.W.Lee
et al.
(2005).
Crystal structures of the main peptidase from the SARS coronavirus inhibited by a substrate-like aza-peptide epoxide.
J Mol Biol,
353,
1137-1151.
PubMed id:
DOI:

|
|
|
Date:
|
|
|
30-Jun-05
|
Release date:
|
25-Oct-05
|
|
|
|
|
|
|
PROCHECK
|

|
|
|
|
|
Headers
|
|
|
|
References
|
|
|
|
|
|

|
|
P0C6X7
(R1AB_CVHSA) -
Replicase polyprotein 1ab from Severe acute respiratory syndrome coronavirus
|
|
|
|
Seq:
Struc:
|
|
|
|
7073 a.a.
306 a.a.
|
|

|

|
|
|
|
|
|
|
|
|
|
|
|
Key: |
 |
PfamA domain |
|
|
 |
Secondary structure |
|
 |
CATH domain |
|
|
|
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.2.1.1.-
- ?????
|
|
|
|
|
|
|
Enzyme class 3:
|
|
E.C.2.1.1.56
- mRNA (guanine-N(7))-methyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L- methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-homocysteine
|
|
|
|
|
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L- methionine
S-adenosyl-L- methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L-homocysteine
S-adenosyl-L-homocysteine
|
|
|
|
|
|
|
|
|
|
Enzyme class 4:
|
|
E.C.2.1.1.57
- methyltransferase cap1.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA + S-adenosyl-L-homocysteine + H+
|
|
|
|
|
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
S-adenosyl-L-methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA
|
+
|
S-adenosyl-L-homocysteine
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 5:
|
|
E.C.2.7.7.48
- RNA-directed Rna polymerase.
|
|
|
|
|
|
|
Reaction:
|
|
RNA(n) + a ribonucleoside 5'-triphosphate = RNA(n+1) + diphosphate
|
|
|
|
|
|
 RNA(n)
RNA(n)
|
+
|
ribonucleoside 5'-triphosphate
|
=
|
RNA(n+1)
|
+
|
 diphosphate
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 6:
|
|
E.C.2.7.7.50
- mRNA guanylyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end diphospho-ribonucleoside in mRNA + GTP + H+ = a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + diphosphate
|
|
|
|
|
|
5'-end diphospho-ribonucleoside in mRNA
|
+
|
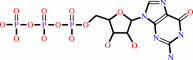 GTP
GTP
|
+
|
H(+)
|
=
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 7:
|
|
E.C.3.1.13.-
- ?????
|
|
|
|
|
|
|
Enzyme class 8:
|
|
E.C.3.4.19.12
- ubiquitinyl hydrolase 1.
|
|
|
|
|
|
|
Reaction:
|
|
Thiol-dependent hydrolysis of ester, thiolester, amide, peptide and isopeptide bonds formed by the C-terminal Gly of ubiquitin (a 76-residue protein attached to proteins as an intracellular targeting signal).
|
|
|
|
|
|
Enzyme class 9:
|
|
E.C.3.4.22.-
- ?????
|
|
|
|
|
|
|
Enzyme class 10:
|
|
E.C.3.4.22.69
- Sars coronavirus main proteinase.
|
|
|
|
|
|
|
Enzyme class 11:
|
|
E.C.3.6.4.12
- Dna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
 ATP
ATP
|
+
|
H2O
|
=
|
 ADP
ADP
|
+
|
 phosphate
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 12:
|
|
E.C.3.6.4.13
- Rna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
ATP
|
+
|
H2O
|
=
|
ADP
|
+
|
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 13:
|
|
E.C.4.6.1.-
- ?????
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Mol Biol
353:1137-1151
(2005)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Crystal structures of the main peptidase from the SARS coronavirus inhibited by a substrate-like aza-peptide epoxide.
|
|
T.W.Lee,
M.M.Cherney,
C.Huitema,
J.Liu,
K.E.James,
J.C.Powers,
L.D.Eltis,
M.N.James.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The main peptidase (M(pro)) from the coronavirus (CoV) causing severe acute
respiratory syndrome (SARS) is one of the most attractive molecular targets for
the development of anti-SARS agents. We report the irreversible inhibition of
SARS-CoV M(pro) by an aza-peptide epoxide (APE; k(inact)/K(i) = 1900(+/-400)
M(-1) s(-1)). The crystal structures of the M(pro):APE complex in the space
groups C2 and P2(1)2(1)2(1) revealed the formation of a covalent bond between
the catalytic Cys145 S(gamma) atom of the peptidase and the epoxide C3 atom of
the inhibitor, substantiating the mode of action of this class of
cysteine-peptidase inhibitors. The aza-peptide component of APE binds in the
substrate-binding regions of M(pro) in a substrate-like manner, with excellent
structural and chemical complementarity. In addition, the crystal structure of
unbound M(pro) in the space group C2 revealed that the "N-fingers"
(N-terminal residues 1 to 7) of both protomers of M(pro) are well defined and
the substrate-binding regions of both protomers are in the catalytically
competent conformation at the crystallization pH of 6.5, contrary to the
previously determined crystal structures of unbound M(pro) in the space group
P2(1).
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 3.
Figure 3. Binding of APE (orange) in the substrate-binding
regions of SARS-CoV Mpro. (a) Stereo view of the outstanding
density in the F[o] -F[c] map for the structures of the Mpro:APE
complex and protomer B of the Click to view the MathML source-
[0?wchp=dGLbVlz-zSkWA]
complex. (b) The corresponding stereo view for protomer A of the
Click to view the MathML source- [0?wchp=dGLbVlz-zSkWA]
complex.
|
|
Figure 7.
Figure 7. Models for each of the four diastereomers of
Cbz-Leu-Phe-AGln-EP-COOEt binding to SARS-CoV Mpro before the
nucleophilic attack by the Cys145 Sg atom of the peptidase. (a)
2S, 3S, (b) 2R, 3R, (c) 2S, 3R, (d) 2R, 3S.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Elsevier:
J Mol Biol
(2005,
353,
1137-1151)
copyright 2005.
|
|
| |
Figures were
selected
by the author.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
C.Niu,
J.Yin,
J.Zhang,
J.C.Vederas,
and
M.N.James
(2008).
Molecular docking identifies the binding of 3-chloropyridine moieties specifically to the S1 pocket of SARS-CoV Mpro.
|
| |
Bioorg Med Chem,
16,
293-302.
|
|
|
|
|
|
|
J.Shi,
J.Sivaraman,
and
J.Song
(2008).
Mechanism for controlling the dimer-monomer switch and coupling dimerization to catalysis of the severe acute respiratory syndrome coronavirus 3C-like protease.
|
| |
J Virol,
82,
4620-4629.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
M.R.Suresh,
P.K.Bhatnagar,
and
D.Das
(2008).
Molecular targets for diagnostics and therapeutics of severe acute respiratory syndrome (SARS-CoV).
|
| |
J Pharm Pharm Sci,
11,
1s.
|
|
|
|
|
|
|
N.Zhong,
S.Zhang,
P.Zou,
J.Chen,
X.Kang,
Z.Li,
C.Liang,
C.Jin,
and
B.Xia
(2008).
Without its N-finger, the main protease of severe acute respiratory syndrome coronavirus can form a novel dimer through its C-terminal domain.
|
| |
J Virol,
82,
4227-4234.
|
|
|
|
|
|
|
R.L.Graham,
J.S.Sparks,
L.D.Eckerle,
A.C.Sims,
and
M.R.Denison
(2008).
SARS coronavirus replicase proteins in pathogenesis.
|
| |
Virus Res,
133,
88.
|
|
|
|
|
|
|
S.Chen,
T.Hu,
J.Zhang,
J.Chen,
K.Chen,
J.Ding,
H.Jiang,
and
X.Shen
(2008).
Mutation of Gly-11 on the dimer interface results in the complete crystallographic dimer dissociation of severe acute respiratory syndrome coronavirus 3C-like protease: crystal structure with molecular dynamics simulations.
|
| |
J Biol Chem,
283,
554-564.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
X.Xue,
H.Yu,
H.Yang,
F.Xue,
Z.Wu,
W.Shen,
J.Li,
Z.Zhou,
Y.Ding,
Q.Zhao,
X.C.Zhang,
M.Liao,
M.Bartlam,
and
Z.Rao
(2008).
Structures of two coronavirus main proteases: implications for substrate binding and antiviral drug design.
|
| |
J Virol,
82,
2515-2527.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
A.K.Ghosh,
K.Xi,
M.E.Johnson,
S.C.Baker,
and
A.D.Mesecar
(2007).
Progress in Anti-SARS Coronavirus Chemistry, Biology and Chemotherapy.
|
| |
Annu Rep Med Chem,
41,
183-196.
|
|
|
|
|
|
|
D.Cuerrier,
T.Moldoveanu,
R.L.Campbell,
J.Kelly,
B.Yoruk,
S.H.Verhelst,
D.Greenbaum,
M.Bogyo,
and
P.L.Davies
(2007).
Development of calpain-specific inactivators by screening of positional scanning epoxide libraries.
|
| |
J Biol Chem,
282,
9600-9611.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
H.P.Chang,
C.Y.Chou,
and
G.G.Chang
(2007).
Reversible unfolding of the severe acute respiratory syndrome coronavirus main protease in guanidinium chloride.
|
| |
Biophys J,
92,
1374-1383.
|
|
|
|
|
|
|
Y.M.Shao,
W.B.Yang,
H.P.Peng,
M.F.Hsu,
K.C.Tsai,
T.H.Kuo,
A.H.Wang,
P.H.Liang,
C.H.Lin,
A.S.Yang,
and
C.H.Wong
(2007).
Structure-based design and synthesis of highly potent SARS-CoV 3CL protease inhibitors.
|
| |
Chembiochem,
8,
1654-1657.
|
|
|
|
|
|
|
E.De Clercq
(2006).
Potential antivirals and antiviral strategies against SARS coronavirus infections.
|
| |
Expert Rev Anti Infect Ther,
4,
291-302.
|
|
|
|
|
|
|
H.Chen,
P.Wei,
C.Huang,
L.Tan,
Y.Liu,
and
L.Lai
(2006).
Only one protomer is active in the dimer of SARS 3C-like proteinase.
|
| |
J Biol Chem,
281,
13894-13898.
|
|
|
|
|
|
|
J.Barrila,
U.Bacha,
and
E.Freire
(2006).
Long-range cooperative interactions modulate dimerization in SARS 3CLpro.
|
| |
Biochemistry,
45,
14908-14916.
|
|
|
|
|
|
|
J.R.Mesters,
J.Tan,
and
R.Hilgenfeld
(2006).
Viral enzymes.
|
| |
Curr Opin Struct Biol,
16,
776-786.
|
|
|
|
|
|
|
M.N.James
(2006).
The peptidases from fungi and viruses.
|
| |
Biol Chem,
387,
1023-1029.
|
|
|
|
|
|
|
S.I.Al-Gharabli,
S.T.Shah,
S.Weik,
M.F.Schmidt,
J.R.Mesters,
D.Kuhn,
G.Klebe,
R.Hilgenfeld,
and
J.Rademann
(2006).
An efficient method for the synthesis of peptide aldehyde libraries employed in the discovery of reversible SARS coronavirus main protease (SARS-CoV Mpro) inhibitors.
|
| |
Chembiochem,
7,
1048-1055.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
code is
shown on the right.
|
|


 Links
Links
































