 |
PDBsum entry 1ls1

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Protein transport
|
PDB id
|
|
|
|
1ls1
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.3.6.5.4
- signal-recognition-particle GTPase.
|
|
|
|
|
|
|
Reaction:
|
|
GTP + H2O = GDP + phosphate + H+
|
|
|
|
|
|
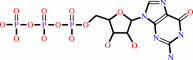 GTP
GTP
|
+
|
H2O
|
=
|
 GDP
GDP
|
+
|
 phosphate
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Mol Biol
320:783-799
(2002)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Structural basis for mobility in the 1.1 A crystal structure of the NG domain of Thermus aquaticus Ffh.
|
|
U.D.Ramirez,
G.Minasov,
P.J.Focia,
R.M.Stroud,
P.Walter,
P.Kuhn,
D.M.Freymann.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The NG domain of the prokaryotic signal recognition protein Ffh is a two-domain
GTPase that comprises part of the prokaryotic signal recognition particle (SRP)
that functions in co-translational targeting of proteins to the membrane. The
interface between the N and G domains includes two highly conserved sequence
motifs and is adjacent in sequence and structure to one of the conserved GTPase
signature motifs. Previous structural studies have shown that the relative
orientation of the two domains is dynamic. The N domain of Ffh has been proposed
to function in regulating the nucleotide-binding interactions of the G domain.
However, biochemical studies suggest a more complex role for the domain in
integrating communication between signal sequence recognition and interaction
with receptor. Here, we report the structure of the apo NG GTPase of Ffh from
Thermus aquaticus refined at 1.10 A resolution. Although the G domain is very
well ordered in this structure, the N domain is less well ordered, reflecting
the dynamic relationship between the two domains previously inferred. We
demonstrate that the anisotropic displacement parameters directly visualize the
underlying mobility between the two domains, and present a detailed structural
analysis of the packing of the residues, including the critical alpha4 helix,
that comprise the interface. Our data allows us to propose a structural
explanation for the functional significance of sequence elements conserved at
the N/G interface.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 3.
Figure 3. Anisotropic temperature factors reflect coherent
motion of the N-domain. The anisotropic ellipsoids of motion at
the 90% probability level for the a-carbon atoms of helices aN1
and aN2 highlight the mobility of the main-chain atoms of the
N-domain. The principal axes of vibration along both of the
helices are largely coherent (  vert,
similar up and down; compare with the ellipsoids near residue
1). The direction of this motion (presumably trapped substrates
in the crystal) is similar to that seen between the apo and GDP
complex crystal structures. vert,
similar up and down; compare with the ellipsoids near residue
1). The direction of this motion (presumably trapped substrates
in the crystal) is similar to that seen between the apo and GDP
complex crystal structures.
|
|
Figure 8.
Figure 8. Conserved residues at the NG interface contribute
to a well-packed interface. Small probe contact dots[8] are
shown at a dot density of 50 dots/Å2. The color of the
dots indicates the types of interactions, with blue dots
indicating wide contacts, greater than 0.25 Å, green
indicating close contacts between 0 Å and 0.25 Å,
yellow indicating small overlaps of less than 0.2 Å, and
purple indicating hydrogen bonds. Hydrogen atoms are included in
the analysis, but are not shown in the Figure. (a) The packing
density around Gly253 and Gly254 (at the center) is especially
tight for glycine residues and thus explains conservation of
these two residues. (b) Leu257 appears to have relatively few
interactions; however, those are with highly conserved residues,
including Leu5, Val43, Leu38, and Leu82. Note the interaction
between Leu5 and Leu257, which involves methyl hydrogen atoms of
the two residues. (c) Interactions with Ser258, which is a
completely conserved residue in the SRP GTPases. Both the
Gly253/Gly254 and Ser258 interactions serve as the "glue" across
the N/G interaction surface.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Elsevier:
J Mol Biol
(2002,
320,
783-799)
copyright 2002.
|
|
| |
Figures were
selected
by the author.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
M.Yang,
X.Zhang,
and
K.Han
(2010).
Molecular dynamics simulation of SRP GTPases: towards an understanding of the complex formation from equilibrium fluctuations.
|
| |
Proteins,
78,
2222-2237.
|
|
|
|
|
|
|
B.H.Kim,
H.Cheng,
and
N.V.Grishin
(2009).
HorA web server to infer homology between proteins using sequence and structural similarity.
|
| |
Nucleic Acids Res,
37,
W532-W538.
|
|
|
|
|
|
|
E.M.Clérico,
J.L.Maki,
and
L.M.Gierasch
(2008).
Use of synthetic signal sequences to explore the protein export machinery.
|
| |
Biopolymers,
90,
307-319.
|
|
|
|
|
|
|
U.D.Ramirez,
P.J.Focia,
and
D.M.Freymann
(2008).
Nucleotide-binding flexibility in ultrahigh-resolution structures of the SRP GTPase Ffh.
|
| |
Acta Crystallogr D Biol Crystallogr,
64,
1043-1053.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
X.Zhang,
S.Kung,
and
S.O.Shan
(2008).
Demonstration of a multistep mechanism for assembly of the SRP x SRP receptor complex: implications for the catalytic role of SRP RNA.
|
| |
J Mol Biol,
381,
581-593.
|
|
|
|
|
|
|
G.Bange,
G.Petzold,
K.Wild,
R.O.Parlitz,
and
I.Sinning
(2007).
The crystal structure of the third signal-recognition particle GTPase FlhF reveals a homodimer with bound GTP.
|
| |
Proc Natl Acad Sci U S A,
104,
13621-13625.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
J.Gawronski-Salerno,
and
D.M.Freymann
(2007).
Structure of the GMPPNP-stabilized NG domain complex of the SRP GTPases Ffh and FtsY.
|
| |
J Struct Biol,
158,
122-128.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
J.Gawronski-Salerno,
J.S.Coon,
P.J.Focia,
and
D.M.Freymann
(2007).
X-ray structure of the T. aquaticus FtsY:GDP complex suggests functional roles for the C-terminal helix of the SRP GTPases.
|
| |
Proteins,
66,
984-995.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
S.Mouilleron,
and
B.Golinelli-Pimpaneau
(2007).
Domain motions of glucosamine-6P synthase: comparison of the anisotropic displacements in the crystals and the catalytic hinge-bending rotation.
|
| |
Protein Sci,
16,
485-493.
|
|
|
|
|
|
|
T.Hainzl,
S.Huang,
and
A.E.Sauer-Eriksson
(2007).
Interaction of signal-recognition particle 54 GTPase domain and signal-recognition particle RNA in the free signal-recognition particle.
|
| |
Proc Natl Acad Sci U S A,
104,
14911-14916.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
H.J.Dong,
S.M.Tao,
Y.Q.Li,
S.H.Chan,
X.L.Shen,
C.X.Wang,
and
W.J.Guan
(2006).
Analysis of the GTPase activity and active sites of the NG domains of FtsY and Ffh from Streptomyces coelicolor.
|
| |
Acta Biochim Biophys Sin (Shanghai),
38,
467-476.
|
|
|
|
|
|
|
U.D.Ramirez,
and
D.M.Freymann
(2006).
Analysis of protein hydration in ultrahigh-resolution structures of the SRP GTPase Ffh.
|
| |
Acta Crystallogr D Biol Crystallogr,
62,
1520-1534.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
B.Zambelli,
M.Stola,
F.Musiani,
K.De Vriendt,
B.Samyn,
B.Devreese,
J.Van Beeumen,
P.Turano,
A.Dikiy,
D.A.Bryant,
and
S.Ciurli
(2005).
UreG, a chaperone in the urease assembly process, is an intrinsically unstructured GTPase that specifically binds Zn2+.
|
| |
J Biol Chem,
280,
4684-4695.
|
|
|
|
|
|
|
J.A.Doudna,
and
R.T.Batey
(2004).
Structural insights into the signal recognition particle.
|
| |
Annu Rev Biochem,
73,
539-557.
|
|
|
|
|
|
|
K.Wild,
K.R.Rosendal,
and
I.Sinning
(2004).
A structural step into the SRP cycle.
|
| |
Mol Microbiol,
53,
357-363.
|
|
|
|
|
|
|
M.J.Bernett,
T.Somasundaram,
and
M.Blaber
(2004).
An atomic resolution structure for human fibroblast growth factor 1.
|
| |
Proteins,
57,
626-634.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
P.F.Egea,
S.O.Shan,
J.Napetschnig,
D.F.Savage,
P.Walter,
and
R.M.Stroud
(2004).
Substrate twinning activates the signal recognition particle and its receptor.
|
| |
Nature,
427,
215-221.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
P.J.Focia,
H.Alam,
T.Lu,
U.D.Ramirez,
and
D.M.Freymann
(2004).
Novel protein and Mg2+ configurations in the Mg2+GDP complex of the SRP GTPase ffh.
|
| |
Proteins,
54,
222-230.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
P.J.Focia,
I.V.Shepotinovskaya,
J.A.Seidler,
and
D.M.Freymann
(2004).
Heterodimeric GTPase core of the SRP targeting complex.
|
| |
Science,
303,
373-377.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
I.V.Shepotinovskaya,
P.J.Focia,
and
D.M.Freymann
(2003).
Crystallization of the GMPPCP complex of the NG domains of Thermus aquaticus Ffh and FtsY.
|
| |
Acta Crystallogr D Biol Crystallogr,
59,
1834-1837.
|
|
|
|
|
|
|
K.Nagai,
C.Oubridge,
A.Kuglstatter,
E.Menichelli,
C.Isel,
and
L.Jovine
(2003).
Structure, function and evolution of the signal recognition particle.
|
| |
EMBO J,
22,
3479-3485.
|
|
|
|
|
|
|
K.R.Rosendal,
K.Wild,
G.Montoya,
and
I.Sinning
(2003).
Crystal structure of the complete core of archaeal signal recognition particle and implications for interdomain communication.
|
| |
Proc Natl Acad Sci U S A,
100,
14701-14706.
|
|
|
PDB codes:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|



 Links
Links













