 |
PDBsum entry 2gop

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
Chains A, B:
E.C.?
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
DOI no:
|
Acta Crystallogr D Biol Crystallogr
63:179-187
(2007)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
The beta-propeller domain of the trilobed protease from Pyrococcus furiosus reveals an open Velcro topology.
|
|
J.Bosch,
T.Tamura,
N.Tamura,
W.Baumeister,
L.O.Essen.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
In the proteolytic pathway of prokaryotic and eukaryotic organisms, proteins
tagged for proteolysis are firstly shredded into smaller peptides by
compartmentalized proteases such as the proteasome complex. Accordingly, a
variety of downstream proteases have evolved to further hydrolyze these peptides
to the level of free amino acids. In the search for such downstream proteases, a
high-molecular-weight protease complex called trilobed protease (TLP) was
recently discovered in the archaeon Pyroccocus furiosus. The crystal structure
of the N-terminal beta-propeller domain of the trilobed protease at 2 A
resolution shows that the trilobed protease utilizes this accessory domain to
control substrate access to the active site. Modelling of the intact TLP monomer
suggests that this protease has an additional side entrance to its active site
as in the DPP-IV or tricorn protease complexes.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 3.
Figure 3 (a) Three-dimensional representation of a hypothetical
model of the intact trilobed protease. Structure prediction of
the  / /
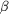 -hydrolase
domain was performed using the LIBELLULA server (Juan et al.,
2003[Juan, D., Grana, O., Pazos, F., Fariselli, P., Casadio, R.
& Valencia, A. (2003). Proteins, 50, 600-608.]) and is shown in
brown. The -propeller
domain is depicted as in the previous figures. Both domains were
fitted using the program TOP (Lu, 2000[Lu, G. (2000). J. Appl.
Cryst. 33, 176-183.]) according to the overall structure of
prolyl oligopeptidase (Fülöp et al.,
1998[Fülöp, V., Bocskei, Z. & Polgar, L. (1998). Cell,
94, 161-170.]). Residues of the active site are shown as green
spheres. (b) Schematic overview of possible substrate pathways
in the monomeric trilobed enzyme. Green arrows represent
substrate access to the active site; the catalytic domain is
represented as a cone-shaped domain and the -propeller
domain is colour coded by individual blades. The first model
represents a diffuse only model, where the pore cannot be
widened, and the second model allows a limited widening of the
central pore; both cases are applicable to the Velcro-type
closures of -propeller
domains. The third and the fourth model result in larger
movements of the -propeller
domain and are limited to non-Velcro-type -propellers,
allowing a larger breathing mechanism for the uptake of larger
substrates. The last model is a combination of the breathing
mechanism and an additional domain movement, allowing much
larger substrates to access the catalytic site via the side
entrance. -hydrolase
domain was performed using the LIBELLULA server (Juan et al.,
2003[Juan, D., Grana, O., Pazos, F., Fariselli, P., Casadio, R.
& Valencia, A. (2003). Proteins, 50, 600-608.]) and is shown in
brown. The -propeller
domain is depicted as in the previous figures. Both domains were
fitted using the program TOP (Lu, 2000[Lu, G. (2000). J. Appl.
Cryst. 33, 176-183.]) according to the overall structure of
prolyl oligopeptidase (Fülöp et al.,
1998[Fülöp, V., Bocskei, Z. & Polgar, L. (1998). Cell,
94, 161-170.]). Residues of the active site are shown as green
spheres. (b) Schematic overview of possible substrate pathways
in the monomeric trilobed enzyme. Green arrows represent
substrate access to the active site; the catalytic domain is
represented as a cone-shaped domain and the -propeller
domain is colour coded by individual blades. The first model
represents a diffuse only model, where the pore cannot be
widened, and the second model allows a limited widening of the
central pore; both cases are applicable to the Velcro-type
closures of -propeller
domains. The third and the fourth model result in larger
movements of the -propeller
domain and are limited to non-Velcro-type -propellers,
allowing a larger breathing mechanism for the uptake of larger
substrates. The last model is a combination of the breathing
mechanism and an additional domain movement, allowing much
larger substrates to access the catalytic site via the side
entrance.
|
|
Figure 4.
Figure 4 Comparison of TLP- with
other -propeller
domains from compartmentalized proteases. (a) Ribbon diagram of
TLP- as
viewed along the sevenfold pseudo-symmetry axis. The nine
aspartate residues forming the negative ring at the entrance to
the channel are highlighted as a ball-and-stick model. (b)
Electrostatic surface representation of TLP- in
top, bottom and cut-open side views. Negative surface charges
are coloured red and positive charges blue. (c) Surface
representation (colour codes: aromatic, green; aliphatic,
yellow; polar, cyan; charged, grey) showing the interior side of
the channel of the superimposed structures of TLP- (I),
POP (II), TRI- 7
(III) and DPP-IV (IV) from the bottom view.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from the IUCr:
Acta Crystallogr D Biol Crystallogr
(2007,
63,
179-187)
copyright 2007.
|
|
| |
Figures were
selected
by the author.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
|


 Links
Links















