 |
PDBsum entry 1a0f

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.2.5.1.18
- glutathione transferase.
|
|
|
|
|
|
|
Reaction:
|
|
RX + glutathione = an S-substituted glutathione + a halide anion + H+
|
|
|
|
|
|
RX
Bound ligand (Het Group name = )
matches with 86.96% similarity
|
+
|
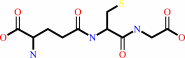 glutathione
glutathione
|
=
|
S-substituted glutathione
|
+
|
halide anion
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Mol Biol
281:135-147
(1998)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Three-dimensional structure of Escherichia coli glutathione S-transferase complexed with glutathione sulfonate: catalytic roles of Cys10 and His106.
|
|
M.Nishida,
S.Harada,
S.Noguchi,
Y.Satow,
H.Inoue,
K.Takahashi.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Cytosolic glutathione S-transferase is a family of multi-functional enzymes
involved in the detoxification of a large variety of xenobiotic and endobiotic
compounds through glutathione conjugation. The three-dimensional structure of
Escherichia coli glutathione S-transferase complexed with glutathione sulfonate,
N-(N-L-gamma-glutamyl-3-sulfo-L-alanyl)-glycine, has been determined by the
multiple isomorphous replacement method and refined to a crystallographic R
factor of 0.183 at 2.1 A resolution.The E. coli enzyme is a globular homodimer
with dimensions of 58 Ax56 Ax52 A. Each subunit, consisting of a polypeptide of
201 amino acid residues, is divided into a smaller N-terminal domain (residues 1
to 80) and a larger C-terminal one (residues 89 to 201). The core of the
N-terminal domain is constructed by a four-stranded beta-sheet and two
alpha-helices, and that of the C-terminal one is constructed by a right-handed
bundle of four alpha-helices. Glutathione sulfonate, a competitive inhibitor
against glutathione, is bound in a cleft between the N and C-terminal domains.
Therefore, the E. coli enzyme conserves overall constructions common to the
eukaryotic enzymes, in its polypeptide fold, dimeric assembly, and
glutathione-binding site. In the case of the eukaryotic enzymes, tyrosine and
serine residues near the N terminus are located in the proximity of the sulfur
atom of the bound glutathione, and are proposed to be catalytically essential.
In the E. coli enzyme, Tyr5 and Ser11 corresponding to these residues are not
involved in the interaction with the inhibitor, although they are located in the
vicinity of catalytic site. Instead, Cys10 N and His106 Nepsilon2 atoms are
hydrogen-bonded to the sulfonate group of the inhibitor. On the basis of this
structural study, Cys10 and His106 are ascribed to the catalytic residues that
are distinctive from the family of the eukaryotic enzymes. We propose that
glutathione S-transferases have diverged from a common origin and acquired
different catalytic apparatuses in the process of evolution.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 5.
Figure 5. Schematic drawing for
interactions between E. coli GST
and glutathione sulfonate. Water
molecules located in the GSH-bind-
ing site are shown as Wat. Hydro-
gen bonds are drawn as broken
lines, and their distances are given
in Å . Residues from the accompa-
nying subunit are indicated by
asterisks.
|
|
Figure 7.
Figure 7. Stereo views of the cata-
lytic site of E. coli GST superposed
on the structures of eukaryotic
theta and alpha GSTs. The ligand
molecules, glutathione sulfonate in
E. coli GST, S-hexyl-glutathione
in theta GST from A. thaliana
(Reinemer et al., 1996), and S-ben-
zyl-glutathione in human GST
alpha 1-1 (Sinning et al., 1993), and
the C
a
atoms of the enzymes are
superposed in the Figure. The
structure for the liganded E. coli
GST is colored red, and those for
the theta (a) and alpha (b) GSTs
are in green and blue, respectively.
Residues from the accompanying
subunit are shown by asterisks.
The ligand molecules are in light
colors. Hydrogen bonds formed in
the catalytic site of E. coli and
alpha are drawn as dotted lines.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Elsevier:
J Mol Biol
(1998,
281,
135-147)
copyright 1998.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
L.Federici,
M.Masulli,
C.Di Ilio,
and
N.Allocati
(2010).
Characterization of the hydrophobic substrate-binding site of the bacterial beta class glutathione transferase from Proteus mirabilis.
|
| |
Protein Eng Des Sel,
23,
743-750.
|
|
|
|
|
|
|
L.Xun,
S.M.Belchik,
R.Xun,
Y.Huang,
H.Zhou,
E.Sanchez,
C.Kang,
and
P.G.Board
(2010).
S-Glutathionyl-(chloro)hydroquinone reductases: a novel class of glutathione transferases.
|
| |
Biochem J,
428,
419-427.
|
|
|
|
|
|
|
T.W.Lin,
P.J.Hsieh,
C.L.Lin,
Y.Y.Fang,
J.X.Yang,
C.C.Tsai,
P.L.Chiang,
C.Y.Pan,
and
Y.T.Chen
(2010).
Label-free detection of protein-protein interactions using a calmodulin-modified nanowire transistor.
|
| |
Proc Natl Acad Sci U S A,
107,
1047-1052.
|
|
|
|
|
|
|
J.K.Takimoto,
K.L.Adams,
Z.Xiang,
and
L.Wang
(2009).
Improving orthogonal tRNA-synthetase recognition for efficient unnatural amino acid incorporation and application in mammalian cells.
|
| |
Mol Biosyst,
5,
931-934.
|
|
|
|
|
|
|
N.Allocati,
L.Federici,
M.Masulli,
and
C.Di Ilio
(2009).
Glutathione transferases in bacteria.
|
| |
FEBS J,
276,
58-75.
|
|
|
|
|
|
|
B.Remmerie,
K.Vandenbroucke,
L.De Smet,
W.Carpentier,
D.De Vos,
J.Stout,
J.Van Beeumen,
and
S.N.Savvides
(2008).
Expression, purification, crystallization and structure determination of two glutathione S-transferase-like proteins from Shewanella oneidensis.
|
| |
Acta Crystallogr Sect F Struct Biol Cryst Commun,
64,
548-553.
|
|
|
|
|
|
|
K.J.Kim,
M.C.Park,
S.J.Choi,
Y.S.Oh,
E.C.Choi,
H.J.Cho,
M.H.Kim,
S.H.Kim,
D.W.Kim,
S.Kim,
and
B.S.Kang
(2008).
Determination of three-dimensional structure and residues of the novel tumor suppressor AIMP3/p18 required for the interaction with ATM.
|
| |
J Biol Chem,
283,
14032-14040.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
N.Allocati,
L.Federici,
M.Masulli,
B.Favaloro,
and
C.Di Ilio
(2008).
Cysteine 10 is critical for the activity of Ochrobactrum anthropi glutathione transferase and its mutation to alanine causes the preferential binding of glutathione to the H-site.
|
| |
Proteins,
71,
16-23.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
S.Conn,
C.Curtin,
A.Bézier,
C.Franco,
and
W.Zhang
(2008).
Purification, molecular cloning, and characterization of glutathione S-transferases (GSTs) from pigmented Vitis vinifera L. cell suspension cultures as putative anthocyanin transport proteins.
|
| |
J Exp Bot,
59,
3621-3634.
|
|
|
|
|
|
|
E.Masai,
Y.Katayama,
and
M.Fukuda
(2007).
Genetic and biochemical investigations on bacterial catabolic pathways for lignin-derived aromatic compounds.
|
| |
Biosci Biotechnol Biochem,
71,
1.
|
|
|
|
|
|
|
E.I.Tocheva,
P.D.Fortin,
L.D.Eltis,
and
M.E.Murphy
(2006).
Structures of ternary complexes of BphK, a bacterial glutathione S-transferase that reductively dechlorinates polychlorinated biphenyl metabolites.
|
| |
J Biol Chem,
281,
30933-30940.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
H.Simader,
M.Hothorn,
and
D.Suck
(2006).
Structures of the interacting domains from yeast glutamyl-tRNA synthetase and tRNA-aminoacylation and nuclear-export cofactor Arc1p reveal a novel function for an old fold.
|
| |
Acta Crystallogr D Biol Crystallogr,
62,
1510-1519.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
J.L.Pan,
and
J.C.Bardwell
(2006).
The origami of thioredoxin-like folds.
|
| |
Protein Sci,
15,
2217-2227.
|
|
|
|
|
|
|
A.M.Hansen,
Y.Gu,
M.Li,
M.Andrykovitch,
D.S.Waugh,
D.J.Jin,
and
X.Ji
(2005).
Structural basis for the function of stringent starvation protein a as a transcription factor.
|
| |
J Biol Chem,
280,
17380-17391.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
J.Li,
Z.Xia,
and
J.Ding
(2005).
Thioredoxin-like domain of human kappa class glutathione transferase reveals sequence homology and structure similarity to the theta class enzyme.
|
| |
Protein Sci,
14,
2361-2369.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
K.P.Lim,
and
W.Hong
(2004).
Human Nischarin/imidazoline receptor antisera-selected protein is targeted to the endosomes by a combined action of a PX domain and a coiled-coil region.
|
| |
J Biol Chem,
279,
54770-54782.
|
|
|
|
|
|
|
C.L.Rife,
J.F.Parsons,
G.Xiao,
G.L.Gilliland,
and
R.N.Armstrong
(2003).
Conserved structural elements in glutathione transferase homologues encoded in the genome of Escherichia coli.
|
| |
Proteins,
53,
777-782.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
A.Kentsis,
R.E.Gordon,
and
K.L.Borden
(2002).
Control of biochemical reactions through supramolecular RING domain self-assembly.
|
| |
Proc Natl Acad Sci U S A,
99,
15404-15409.
|
|
|
|
|
|
|
A.M.Caccuri,
G.Antonini,
N.Allocati,
C.Di Ilio,
F.De Maria,
F.Innocenti,
M.W.Parker,
M.Masulli,
M.Lo Bello,
P.Turella,
G.Federici,
and
G.Ricci
(2002).
GSTB1-1 from Proteus mirabilis: a snapshot of an enzyme in the evolutionary pathway from a redox enzyme to a conjugating enzyme.
|
| |
J Biol Chem,
277,
18777-18784.
|
|
|
|
|
|
|
L.Bousset,
H.Belrhali,
J.Janin,
R.Melki,
and
S.Morera
(2001).
Structure of the globular region of the prion protein Ure2 from the yeast Saccharomyces cerevisiae.
|
| |
Structure,
9,
39-46.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
T.C.Umland,
K.L.Taylor,
S.Rhee,
R.B.Wickner,
and
D.R.Davies
(2001).
The crystal structure of the nitrogen regulation fragment of the yeast prion protein Ure2p.
|
| |
Proc Natl Acad Sci U S A,
98,
1459-1464.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
J.U.Flanagan,
J.Rossjohn,
M.W.Parker,
P.G.Board,
and
G.Chelvanayagam
(1999).
Mutagenic analysis of conserved arginine residues in and around the novel sulfate binding pocket of the human Theta class glutathione transferase T2-2.
|
| |
Protein Sci,
8,
2205-2212.
|
|
|
|
|
|
|
Van Hop D,
A.Gaikwad,
B.S.Yadav,
M.K.Reddy,
S.Sopory,
and
S.K.Mukherjee
(1999).
Suppression of pea nuclear topoisomerase I enzyme activity by pea PCNA.
|
| |
Plant J,
19,
153-162.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
code is
shown on the right.
|
|


 Links
Links











